
The retina is the innermost of the three coats of the eye. This layer is responsible for converting relevant information from the image of the external environment into neural impulses that are transmitted to the brain. Broadly retina consists of two primary layers: an inner neurosensory retina and retinal pigment epithelium (RPE). 1
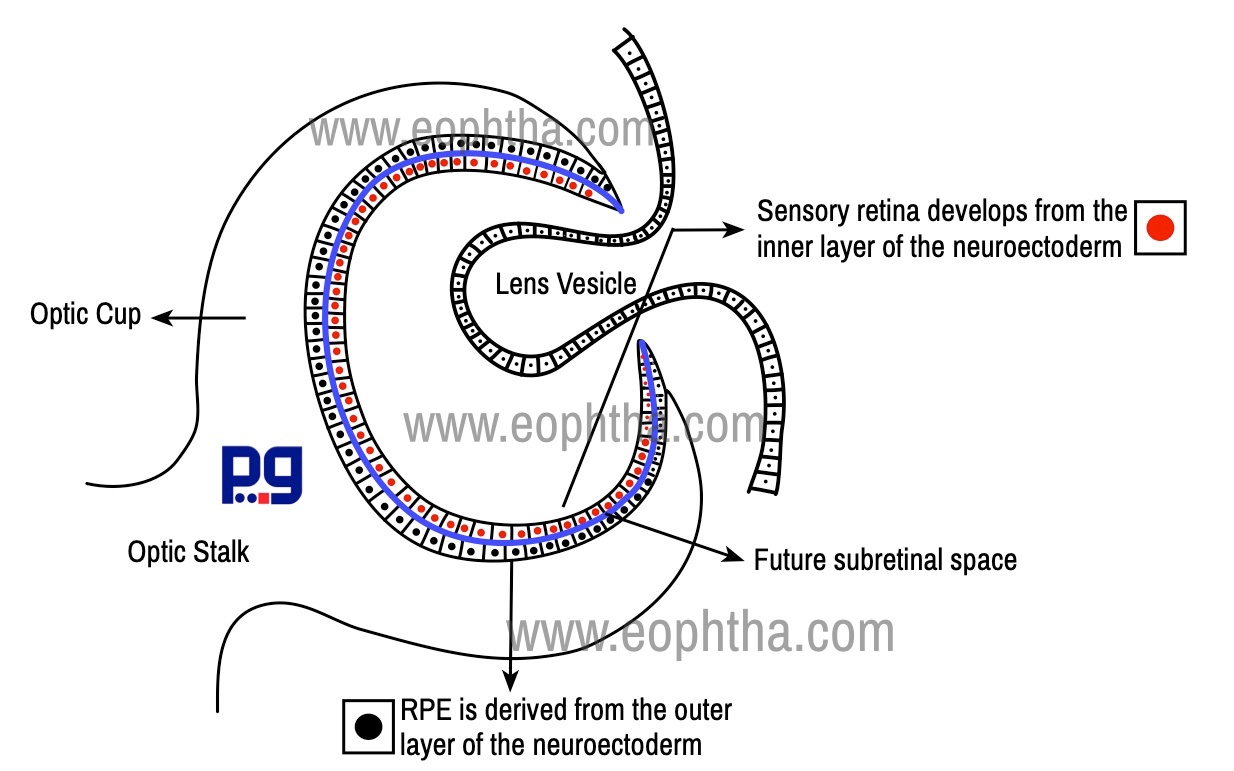
Sensory retina develops from the inner layer of the neuroectoderm, whereas RPE is derived from the outer layer of the neuroectoderm. The RPE is continuous anteriorly with the pigment epithelium of the ciliary body. Sensory retina extends from the optic disc to ora serrata, where it is continuous with non-pigmented ciliary epithelium.1
Between the neural retina and RPE, there is a potential space known as subretinal space. The adhesion between neural retina and RPE is relatively weak. The neural retina is firmly attached at its anterior termination, the ora serrata, and at the margins of the optic nerve head. The ora serrata is located 4-6 mm behind ciliary epithelium and corresponds to the insertion of medial and lateral rectus muscles. Ora serrata has a smooth appearance temporally but serrated nasally.
The retina is bound externally by Bruch's membrane and on its internal aspect by the vitreous. It is continuous with the optic nerve posteriorly, the site of exit of ganglion cell axons from the eye.
The topography of the Retina:
Human retina measures 32 mm from ora to ora along the horizontal meridian and the total surface area of the retina in each eye is approximately 1,100square mm.2 The average thickness of retina is 200 micrometers, it is slightly thicker near optic nerve head and near the macula and gradually thins out at the ora serrata and fovea.3 Broadly the retinal surface can be divided into the following areas:
Area centralis (commonly known as the posterior pole or central retina)
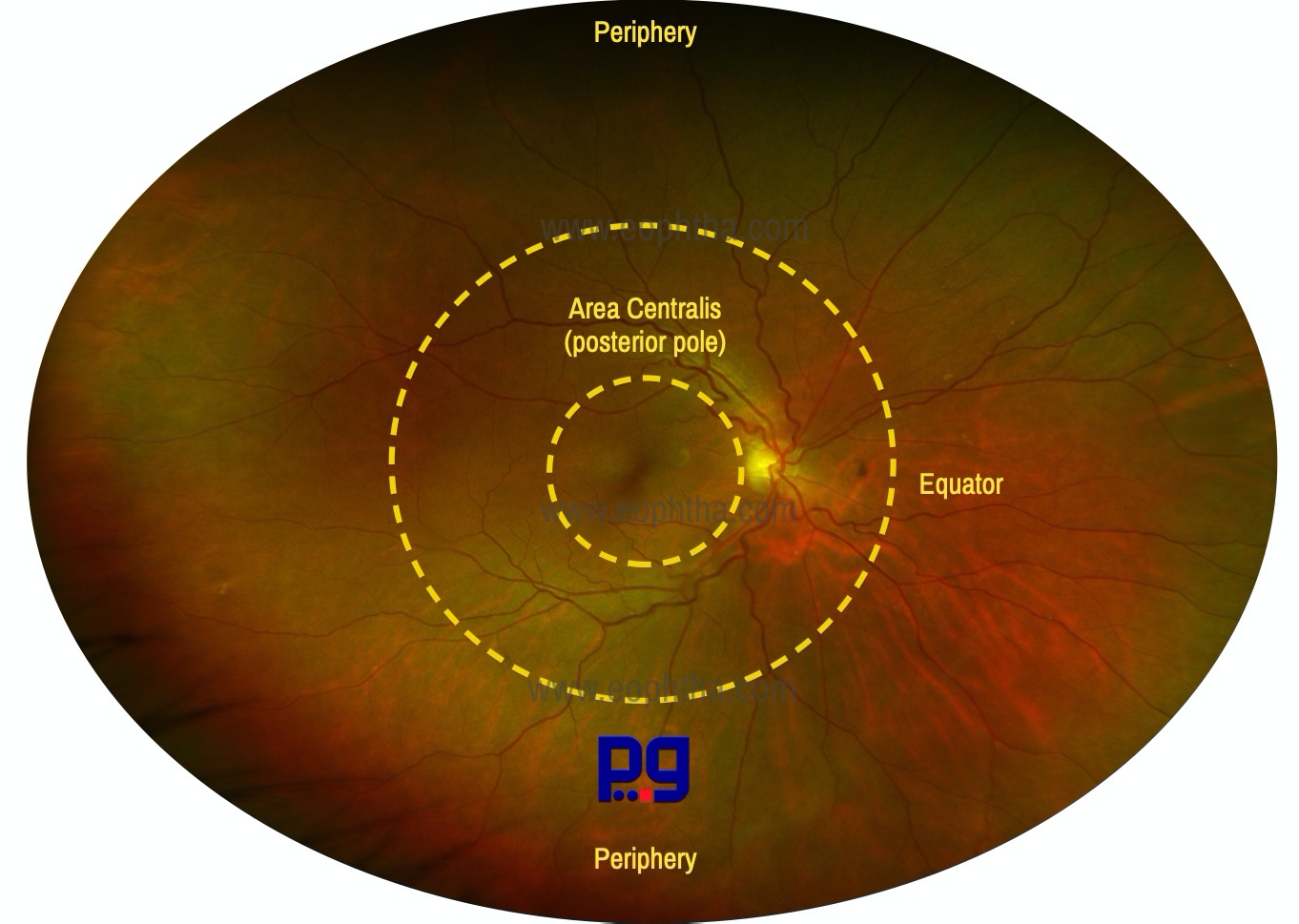
Area centralis is a 5–6 mm diameter circular zone of the retina. It is situated between the superior and inferior temporal arteries and contains
Macula lutea
(Latin Macula means spot; Lutea means yellow) is a 5.5 mm diameter area situated in area centralis, 3 mm lateral to the optic disk. It is partly yellow as a result of yellow xanthophyll carotenoid pigments (zeaxanthin and lutein) in the cone axons. This pigment is believed to act as a short wave-length filter protecting against UV irradiation.
Macula Lutea
Diameter: 5.5 mm
Situation: about 2DD temporal to the disc. (1DD = 1.5 mm)
Colour: Yellow (Due to presence of carotenoid pigment XANTHOPHYLL in the ganglion & bipolar cells)
Macula corresponds to the 150 of the visual fields.
The macula is responsible for photopic vision & color vision is its primary function.
FoveaCentralis (Latin: fovea means small pit or depression) is 1.5mm in diameter. The fovea is fully developed only at the age of 4 years.2 It is located at the posterior pole of the globe, 4 mm temporal to the optic disc & about 0.8 mm below the horizontal meridian. It represents 50 of the visual field. It has an average thickness of 0.25 mm. At the center of the fovea, the layers of the retina are thinner so that central concave indentation, foveola is produced. The downward slopping border of fovea which meets the floor of the foveal pit is known as clivus. The foveal retina is avascular and relies on the choriocapillaris for nutritional support.
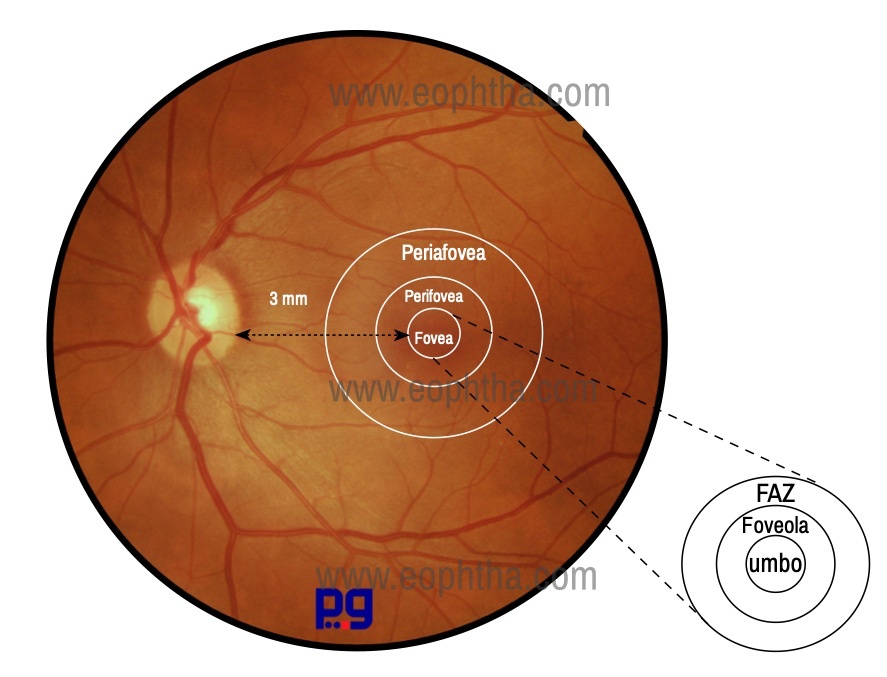
Fovea centralis or Foveola which measures 0.35mm in diameter and cone photoreceptors are concentrated here at a maximum density to the exclusion of rods, the reason why this area represents the area of the highest visual acuity in the retina. Tiny depression in the very center of foveola is known as umbo, which corresponds to the ophthalmoscopically visible foveolar reflex, seen in most of the normal eyes.
|
Macula |
Fovea |
Foveola |
|
Diameter: 5.5mm Represents 150 of the visual field |
Diameter: 1.5mm Thickness: 0.25mm Represents 50 of the visual field |
Diameter: 0.35 mm Thickness: 0.13 mm Represents 10 of the visual field |
Optic Disc The optic disc lies a 3 mm medial to the center of the macula (fovea). There are no normal retinal layers in this zone (blind spot) as ganglion cell axons from the retina pierce the sclera to enter the optic nerve. This pale pink/whitish area is 1.8 mm (1.86 × 1.75 mm)in diameter with a slightly raised rim.2 The central retinal vessels emerge at the center of the optic disk, pass over the rim, and radiate out to supply the retina. The vein usually lies lateral to the artery.

Equator
The equator of the fundus is situated approximately 14 to 15 mm from the limbus. Clinically one of the easiest ways to locate equator of the fundus is to find the vortex veins. The choroidal veins drain into a vortex vein in each quadrant of the eye. Usually there are four vortex veins (one each quadrant), but the number may vary from 8 to 10. The vortex veins traverse the eyeball obliquely through scleral canals for approximately 4 mm and exit the globe posterior to the equator. Some of these vortex veins show dilatations of varying sizes and shapes and are called vortex vein ampullae. Basically, ampulla means the dilated sac of the vortex vein. The equator can be compared with an imaginary circle drawn through the ampullae of the vortex veins. A pigmented crescent or choroidal pigmentation adjacent to a vortex vein ampulla is not uncommon. There are reports that vortex vein varices have been confused with a possible choroidal mass.

Peripheral Retina
The peripheral retina is the remainder of the retina. The distance from the optic disk to the ora serrata is 23–24 mm on the temporal aspect and approximately 18.5 mm on the nasal aspect.6 The peripheral retina is the thinnest (110–140 µm in thickness) and the retina becomes thicker as it progresses towards the posterior pole.
The ora serrata represents the anterior limit of the neural retina. The retina is strongly adherent to the retinal pigment epithelium at the ora serrate, the reason retinal detachment rarely extends anterior to the ora serrata. Ora serrata has a scalloped appearance and is serrated much more nasally than temporally. The width of ora serrate is 2 mm temporally and it is 1 mm wide nasally. When we compare the extension of ora serrata, it is situated 6 mm from the nasal limbus whereas, from the temporal limbus, this distance is 7 mm. Thus, ora serrata is slightly more anterior nasally. Externally, ora serrata corresponds to the insertion of the medial and lateral rectus muscles.
There are two unique structures that can be seen in this region as ora serrata coalesces with pars plana. The rounded extensions of the pars plana at the ora serrata are called ora bays and “teeth-like” extensions of the retina between ora bays are known as dentate processes. There are approximately 20 to 30 dentate processes in each eye.
The ora serrata represents a watershed zone between the anterior and posterior vascular systems and only one layer of blood vessels is present. 1 This may in part explain why peripheral retinal degeneration is relatively common here. The peripheral retina in the region of the ora serrata is markedly attenuated and retina thins to 100 µm here.
Cellular Structures in Retina:
The neurosensory retina is mainly made up of three groups of neurons: photoreceptors, bipolar cells, and ganglion cells. Other important neurons like amacrine cells, horizontal cells have supporting roles. The photoreceptor cells, bipolar cells, and ganglion cells carry the neural signal in a three-step pathway through the retina. Photoreceptors are sensory receptors. Bipolar cells are first-order cells and ganglion cells form second-order neurons. In this section, we will discuss the anatomy of the cells present in the retina.

|
Cell type |
Estimated number /Each Eye |
|
Rod |
approximately 110 to 125 million 2,6 |
|
Cones |
approximately 6.3 to 6.8 million 2,6 |
|
Bipolar cells |
approximately 35.7 million 6 |
|
Ganglion cells |
approximately 1.2 million 6 |
|
RPE cells |
approximately 3.5 million1 |
|
Müller cells |
approximately 10 million |
Retinal Pigment Epithelial Cells (RPE cells):
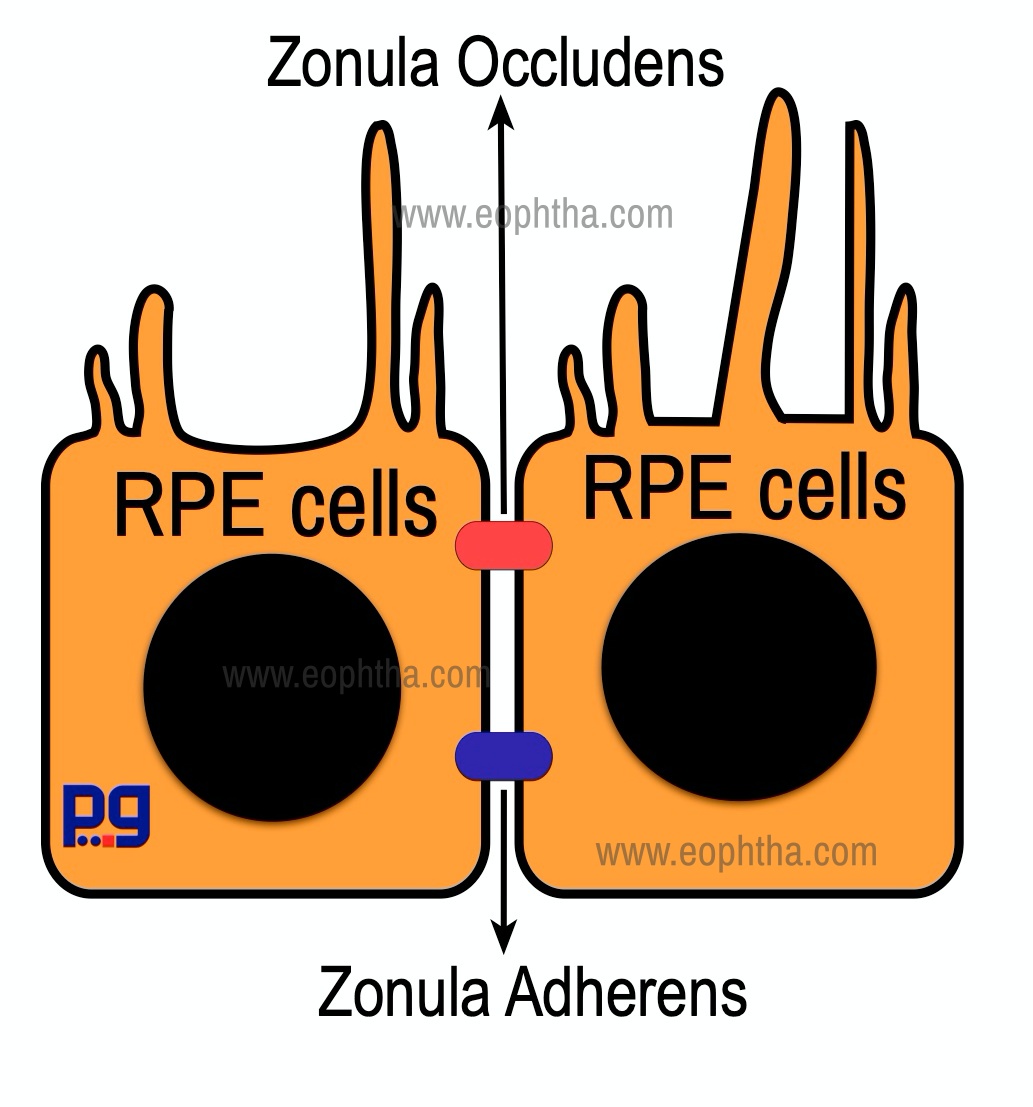
RPE cell is a hexagonal shaped cuboidal cell, situated behind the photoreceptors. RPE cells are connected with each other along their sides and form a single layer that lies between Bruch's membrane and the neurosensory retina. The apical surface of the RPE cell has microvilli that surround the tip of the photoreceptor. The basal part of each RPE cell is attached to the basement membrane of Bruch's membrane by fibronectin, laminin, type IV collagen, and other proteins. Zonula occludens and Zonula adherens play an important role in adhesion of the adjacent RPE cells. Zonula occludens is situated near the apical portion of adjacent RPE cells. Zonula occludens is a tight junction, prevents the passage of molecules, ions, and forms an outer blood-ocular barrier. Zonula adherensdoes not participate in the formation of the blood-ocular barrier, but forms structural adhesions between adjacent RPE cells and retain the monolayer pattern of RPE cells.
These cells vary in size and function in different parts of the retina. For example, RPE cells are taller in the macula, and in the periphery, they are relatively flat. RPE cells have the highest density in the fovea and the density progressively decreases towards the periphery.
The primary function of the RPE cells are:
- Absorption of light passing through the retina
- Regeneration of visual pigments
- Formation of outer blood-ocular barrier
- Maintenance of subretinal space including fluid & electrolyte balance (see RPE pump)
- Phagocytosis of the outer segment of photoreceptor
- Healing and scar tissue formation
RPE cells contain mainly two pigments melanin and lipofuscin. The amount of pigmentation varies from cells to cells, area to area of the retina. It is the variable presence of melanin inside RPE cells that contributes to the typical patterned fundus appearance. Melanin, accumulated in melanosomes, is responsible for the absorption of most damaging wavelengths of visible light.
|
Variation in pigmentation of RPE cells |
|
The brown color of the RPE is imparted by its melanin granules. The highest concentration of pigment is found in the peripheral retina, the lowest in the macular area. With age, RPE gradually loses melanin granules, possibly related to effects of photooxidation. Each RPE cells show variation in pigmentations, the reason for characteristic coarse, granular appearance of fundus. Pigmentation of RPE cells usually increases in periphery of retina with old ages. |
Photoreceptors:
Photoreceptors are the special sense cells which contain photopigments and can absorb photons of light. There are two types of photoreceptors, the rods, and the cones. Rods are responsible for vision in dim light or dark, sensing contrast, brightness, motion and usually produce images of black and white shades. Cones are responsible for vision in bright light, fine resolution, spatial resolution, and color vision. The density of rods and cones varies in different parts of the retina. Rods are absent at fovea, increases in number towards the periphery and then again decreasing in number towards the extreme periphery. The density of cone increases nearer the macula and the foveal region contains only cones.
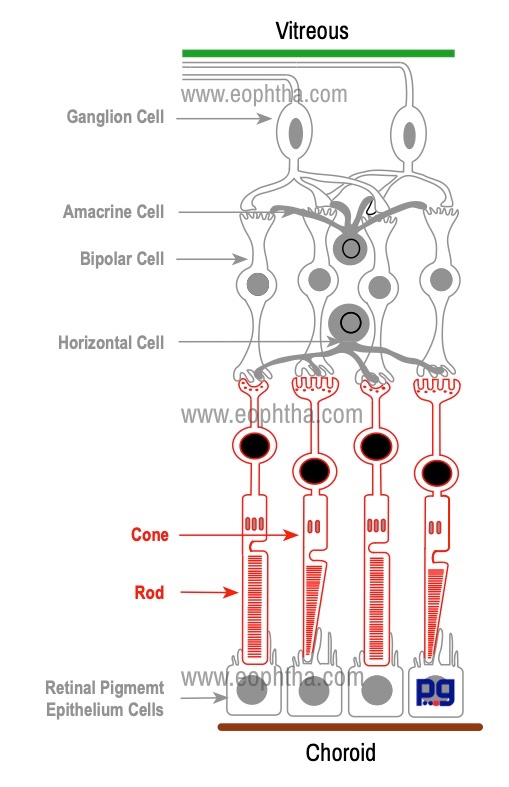
|
Part of the Photoreceptor |
Layer of Retina |
|
Photosensitive part of Rods & Cones |
Photoreceptor Layer/ Layer of Rods & Cones |
|
Separation of inner and outer segments from the cell body |
Outer Limiting Membrane |
|
Nucleus /Cell bodies of Rods & Cones |
Outer Nuclear Layer |
|
Inner processes of Rods & Cones |
Outer Plexiform Layer |
Each photoreceptor is composed of several parts:
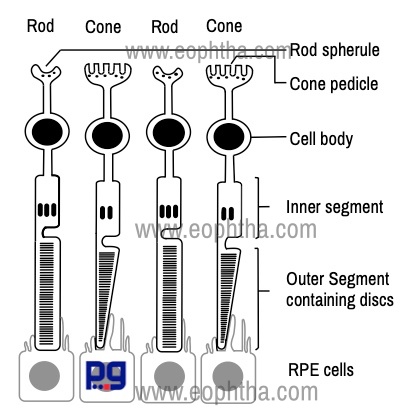
1. Outer segment: outer segment is made up of a stack of membranous discs (600 to 1000 per rod). Each disc is a flattened membrane. These discs are stacked on top of one another and are enclosed in a single membrane. The membrane of the discs contains the visual pigment molecule, which is responsible for the conversion of light into a neural signal. The name of the visual pigment molecule is rhodopsin (maximal spectral sensitivity 496 nm). The discs are produced at the base of the outer segment (towards cilium) and reach the tip of the outer segment which is enclosed by microvilli of RPE cells. Each disc is shed and subsequently phagocytosed by the RPE cells.
|
Parts of a Photoreceptor |
Contents |
|
Outer segment |
Contains visual pigment converts light into a neural signal |
|
Cilium |
Connecting Stalk- joins the outer segment with the inner segment |
|
Inner Segment |
Contains mitochondria, endoplasmic reticulum and Golgi apparatus |
|
Outer Fiber |
Part from inner segment to cell body |
|
Cell body |
The part which contains nucleus |
|
Inner Fiber |
Axonal process - synapse with bipolar and horizontal cells. |
2. Cilium: Cilium is the connecting stalk that joins the outer segment with the inner segment.
3. Inner segment: The innersegment of a photoreceptor can be further divided into two parts: the ellipsoid and the myoid. The ellipsoid is the part nearer to the outer segment. The ellipsoid part contains numerous mitochondria. The myoid, part closer to the cell body, contains other cellular organelles, such as the endoplasmic reticulum and Golgi apparatus.
4. Outer fiber: Outer fiber extends from the inner segment to the cell body.
5. Cell body: Cell body contains the nucleus.
6. Inner fiber:Inner fiber is an axonal process that runs from the cell body to end in specialized synaptic terminals that contain synaptic vesicles. In the case of rods, this synaptic terminal is pear-shaped and called rod spherule. The internal surface of the spherule is invaginated to form a synaptic complex that also contains bipolar dendrites and horizontal cell processes. In a cone, this synaptic terminal is called a cone pedicle.
|
Rod |
Cone |
|
Total number of rods in the human retina has been estimated to be about 110 to 125 million |
Total number of cones in the human retina has been estimated to be about 6.3 to 6.8 million |
|
Rods are responsible for sensing contrast, brightness, and motion |
Cones are responsible for fine resolution, spatial resolution and color vision |
|
Rods are absent at fovea |
Cones are most dense at fovea |
|
The number of rods increases towards the periphery |
Number of cones decreases towards periphery |
|
The outer segment is rod-shaped |
The outer segment is cone-shaped |
|
Only one type, so only monochromatic vision |
Three types (red, green, blue), provides color vision |
|
Contains Rhodopsin |
Contain Photopsin |
|
Regenerative power is high |
Regenerative power is relatively low |
|
Smaller in Size |
Relatively Larger |
|
Slower response speed |
Faster response |
Bipolar Cells:
A bipolar cell is a second-order neuron in the visual pathway which transmits signals from photoreceptors to ganglion cells. Their cell bodies lie in the inner nuclear layer and lie parallel to the photoreceptors. Their single or multiple dendrites synapse principally with photoreceptors (and also with horizontal cells), while their single axon synapses with ganglion and amacrine cells. Bipolar cells have been subdivided into nine morphological subtypes. Principal types of bipolar cells are:
- Rod Bipolar Cells: connect several rods to one to four ganglion cells.
- Flat or Diffuse Bipolar Cells: connect many cones with many ganglion cells
- Midget Bipolar Cells: connect a single cone with a single ganglion cells
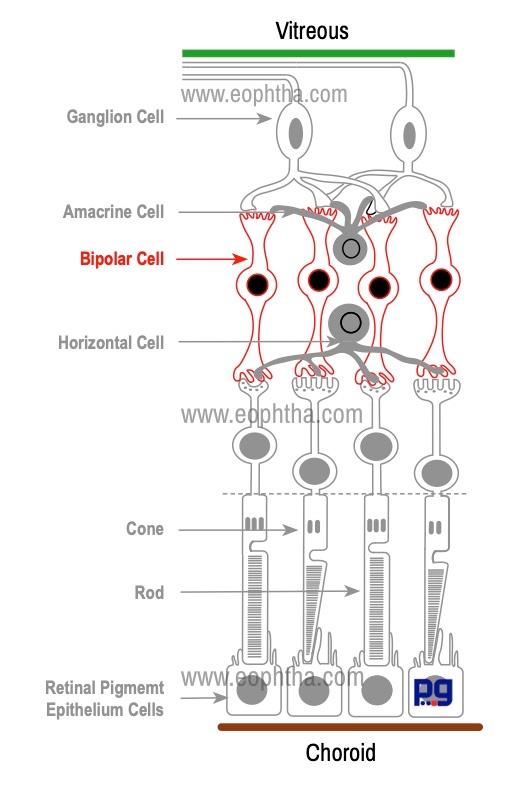
Bipolar cells play a crucial role in transmitting stimuli in the visual pathway. For example, high sensitivity of in foveal region can be explained with the arrangement of photoreceptor and bipolar cells. In this region, the ratio of cones: bipolar cells: ganglion cells is 1: 1: 1, whereas one bipolar cell receives stimuli from up to 50–100 rods in the peripheral retina.
|
Retinal Layer |
Content |
|
Outer Nuclear Layer |
Rods, Cones |
|
Inner Nuclear Layer |
Horizontal cells, bipolar cells, amacrine cells, Müller cells |
|
Ganglion cell layer |
Ganglion cells, displaced amacrine cells |
|
Outer plexiform layer |
Synapses between photoreceptor, bipolar, and horizontal cells |
|
Inner plexiform layer |
Synapses between bipolar, amacrine, and ganglion cells |
Ganglion Cells:
Ganglion cell is the third-order neurons in the visual pathway. The reason for the name " ganglion cell" is their resemblance to cells in nervous ganglia. Usually, they form a single layer in peripheral retina and form as many as 10 layers in the macula.
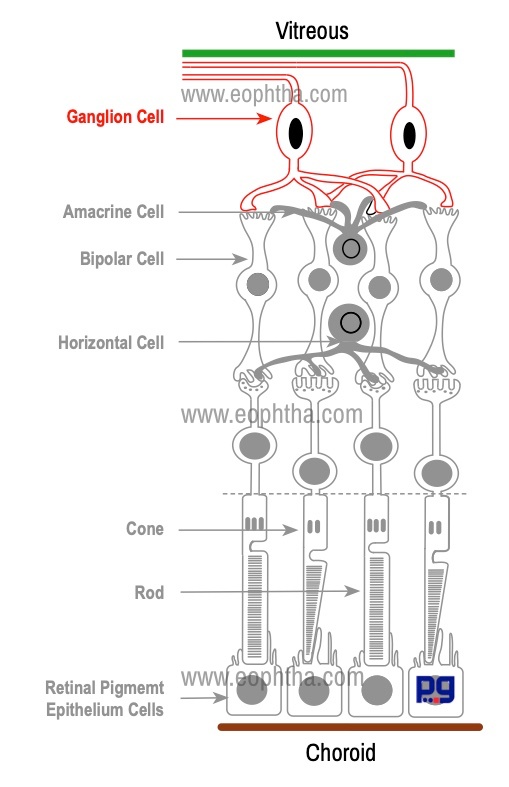
The dendrites of ganglion cells synapse with the axons of bipolar cells and amacrine cells. Each ganglion cell has a single axon and these nonmyelinated axons form the nerve fiber layer on the innermost surface of the retina. These axons make 90 degrees turn to reach the inner surface of the retina and leave the eye as the optic nerve. While coming out from the eye through the lamina cribrosa, these axons become myelinated with oligodendrocytes.
Horizontal Cells:
Horizontal cells lie parallel to the retinal surface. They are situated close the synaptic terminal processes of Photoreceptors. Horizontal cells are multipolar cells with one long axon and several short dendrites with branching terminals. Horizontal cells synapse with photoreceptors, bipolar cells, and other horizontal cells and terminate in the outer plexiform layer.
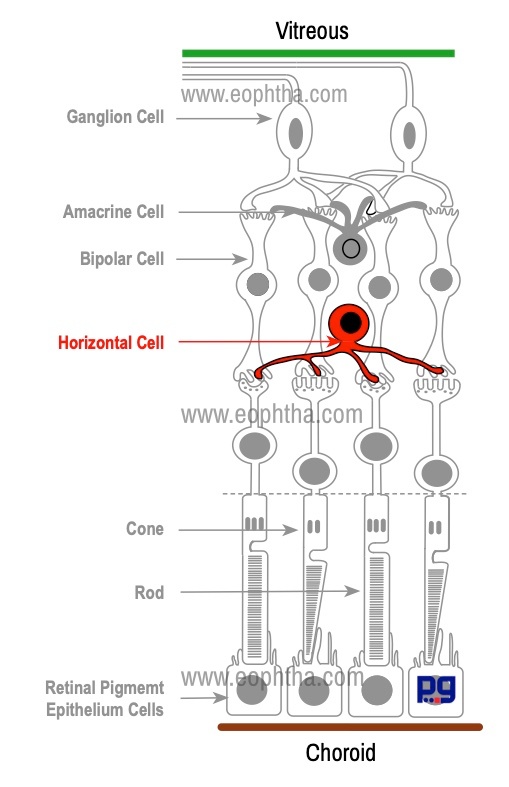
Horizontal cells use the inhibitory neurotransmitter GABA and functions by inhibitory feedback to photoreceptor cells and bipolar cells.
Amacrine Cells:
Amacrine cells (Amacrine: a cell or process that lack long fibrous process) are so named because it was believed to lack axons. They are situated close to the ganglion cells. There are at least 40 types of amacrine cell. They synapse with each other, dendrites of ganglion cells and axons of bipolar cells. Amacrine cells are found in the inner plexiform layer; few displaced amacrine cells are found in the ganglion cell layer
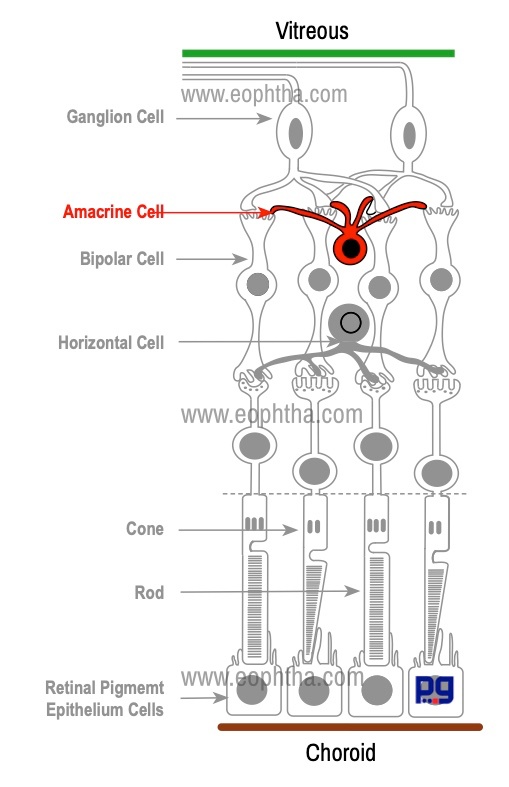
|
Cell type |
Connection |
|
Photoreceptors |
Synapse with bipolar cells and also with horizontal cells |
|
Bipolar cells |
Their single or multiple dendrites synapse principally with photoreceptors (and also with horizontal cells), while their single axon synapses with ganglion and amacrine cells. |
|
Ganglion Cells |
Synapse with the axons of bipolar cells and amacrine cells |
|
Horizontal cells |
Synapse with photoreceptors, bipolar cells, and other horizontal cells. |
|
Amacrine cells |
Synapse with each other, dendrites of ganglion cells and axons of bipolar cells. |
Glial cells of Retina:
The glial cells are specialized cells that surround the neurons and provide support for and insulation between them. Three basic types of glial cell are found in the human retina, Muller cells, astroglia and microglia.
Müller Cells
Müller cells form the major space-occupying tissue of the retina. They are the principal supporting glial cells of the retina. They provide structural as well as nutritional support to the retina. They have a radial orientation and virtually fill in most of the space of the retina not occupied by the neurons. They extend from vitreous to the inner segment of the photoreceptors.
Müller cells perform the following functions:
- They supply end products of anaerobic metabolism to the nerve cells of retina (the breakdown of glycogen) to fuel aerobic metabolism. Müller cells help to nourish and maintain the outer retina, which lacks a direct blood supply
- They clean neural waste products
- Müller cells protect neurons from exposure to excess neurotransmitters
- Müller cells synthesize retinoic acid which helps in the development of the eye
- Müller cells plays a crucial role in homeostasis and neuroprotection
Astrocytes
Astrocytes are star-shaped fibrous cells usually present in the nerve fiber and ganglion cell layer. Retinal astrocytes have processes to insulate retinal vessels and axons.
Layers of Retina:
The retina consists of 10 layers which include nine layers within the sensory retina and retinal pigment epithelium. Embryologically RPE is developed from the outer layer of the optic cup and photoreceptors are developed from the inner layer of the cup and there is a potential space between RPE and sensory retina, known subretinal space. Absence of specialized molecules like laminin and fibronectin, lack of junctional complexes between RPE cells and photoreceptors are responsible for this loose attachment of RPE cells to the photoreceptors. In retinal detachment, the sensory retina detaches from retinal pigment epithelium (RPE). RPE remains firmly attached to the choroid during retinal detachment.
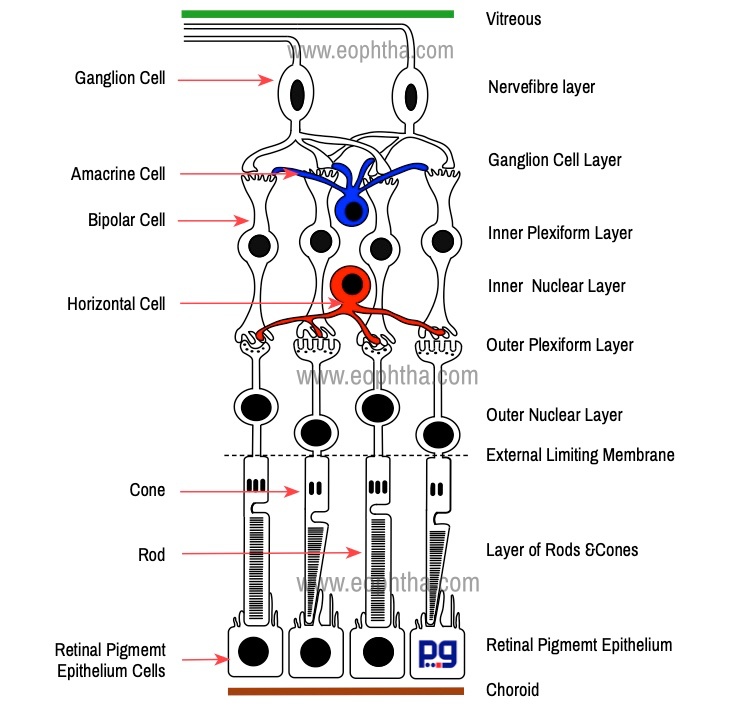
As we have discussed in previous sections retinal cells are arranged in a highly organized way. Cellular components in the retina are organized mainly in five layers: three layers of nucleus/cell bodies and two layers of synapses.
Broadly there are ten layers in Retina: 1. Retinal pigment epithelium 2. Layer of Rods & Cones 3. External limiting membrane 4. Outer nuclear layer 5. Outer plexiform layer 6. Inner nuclear layer 7. Inner plexiform layer 8. Ganglion cell layer 9. Nerve fiber layer 10. Internal limiting membrane. This ten-layered structure was first proposed by Cajal (1888).
Retinal pigment epithelium (RPE):
This is a single cell layer made up of hexagonal shaped RPE cells and lies between Bruch's membrane and the neurosensory retina. There is a potential space between RPE and sensory retina, known as subretinal space. Absence of specialized molecules like laminin and fibronectin, lack of junctional complexes between RPE cells and photoreceptors are responsible for this loose attachment of RPE cells to the photoreceptors. The RPE extends from the optic nerve to the ora serrata and is continuous with the ciliary body pigment epithelium.
Photoreceptors Layer (Layer of rods and cones):
The photoreceptor layer contains only photosensitive part of rods and cones whereas cell bodies and inner processes of them lies in outer nuclear and outer plexiform layers. The central fovea has no rods and foveal region is densely packed with cones (2, 00,000 cones per square millimeter). Outside the fovea, this density of cones gradually decreases. This structural variation results from the centripetal migration of the first order & centrifugal lateral displacement of second & third-order neurons during fetal maturation which occurs 3 months before & 3 months after term.
External Limiting Membrane:
The external limiting membrane is not a true membrane, but a discrete “wire netting” layer. It is composed of zonula adherens junctions between photoreceptor cells and between photoreceptors and Müller cells. Through the fenestrations of the band formed by zonula adherens processes of the rods and cones pass.
This layer often acts as a metabolic barrier restricting the passage of some large molecules.
Outer Nuclear Layer:
The outer nuclear layer consists of rod and cone nuclei. In the peripheral retina, the outer nuclear layer consists of 3 to 4 layers of rod nuclei and a single layer of cone nuclei. It is thickest in the fovea, where it contains approximately 10 layers of cone nuclei
Outer Plexiform Layer:
The word “plexiform” means something related to plexus. The outer plexiform layer is the layer of processes and synapses, which lies in between two nuclear layers (nuclei of various retinal cells). This layer contains synapse and network of branching processes of horizontal cells and bipolar cells along with inner fibers of photoreceptors.
Inner Nuclear Layer:
This layer contains nuclei of horizontal cells, various types of bipolar cells, amacrine cells and Müller cells
Inner Plexiform Layer:
This layer contains processes and synapse of the bipolar, amacrine and ganglion cells. Another unique feature of this layer is that it contains the synapse between the second-order and third-order neuron in the visual pathway
Ganglion Cell Layer:
This is the innermost layer of cell nuclei. In peripheral retina, the ganglion cell layer consists of only a single layer of cells whereas in central retina they form a layer comprise of up to 10 cells. The ganglion cells send a single axon which eventually forms the optic nerve. Thus, ganglion cell is the only layer that is continuous across the optic nerve head.
Nerve Fibre Layer:
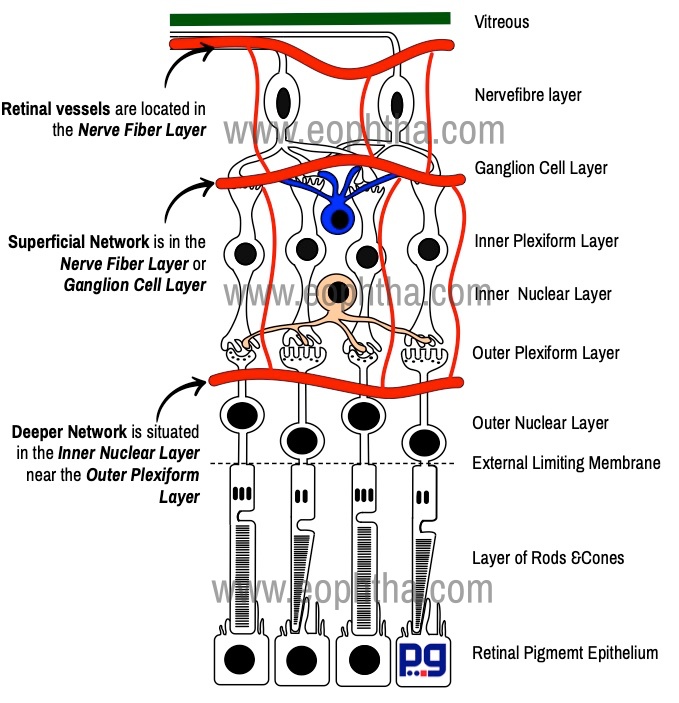
This layer contains axons of the ganglion cells which run parallel to the surface of the retina and emerge out as the optic nerve. These axons or fibers have been grouped according to the area. The group of axons that radiate to the disc from the macular area is called the papillomacular bundle. The temporal retinal raphe is a horizontal boundary separating the superior and inferior retinal nerve fiber bundles in the temporal retina. The retinal vessels are located mainly in the nerve fiber layer (and in the inner nuclear layer). Müller cells play an important role in ensheathing vessels and nerve fibers.
Inner Limiting Membrane:
The inner limiting membrane is the innermost layer of the retina. This layer is made up of extensions of Müller cells covered by a basement membrane.
|
Name of the layers |
Content |
|
RPE |
Retinal pigment epithelial cells |
|
Layer of Photoreceptor |
Inner and Outer segments of Photoreceptor |
|
External limiting membrane |
Made of zonulae adherens between Müller cells and photoreceptor inner segments |
|
Outer nuclear layer |
The photoreceptor cell bodies |
|
Outer plexiform layer |
Synapses between photoreceptor, bipolar, and horizontal cells |
|
Inner nuclear layer |
Horizontal, bipolar, and amacrine cell bodies; Müller cell nuclei |
|
Inner plexiform layer |
Synapses between bipolar, amacrine, and ganglion cells |
|
Ganglion cell layer |
Ganglion cells and displaced amacrine cells |
|
Nerve fiber layer |
Ganglion cell axons traveling towards the optic nerve |
|
Internal limiting membrane |
Formed by Müller cell end feet |
Cellular Arrangements in Retina
The cellular arrangements in the form of layers vary according to the topography of the retina. The foveola, the most sensitive part of the fovea is blue cone and rod free. Foveola largely consists of red, green cones & their nuclei covered by a thin internal limiting membrane. All other retinal layers are absent in this region. Henle’s layer is the foveal portion of the outer plexiform layer of the retina. This layer is characterized by oblique arrangements of the cone axons. In contrast to thevertical orientation of the nerve fibers in the extrafoveal retina, the nerve fibers in this layer course obliquely & actually run parallel to the retinal surface. Also, in the central retina, every cone is interconnected with a single bipolar cell, which in turn synapses with a single ganglion cell. This provides a direct one-to-one relationship between the first three visual neurons.
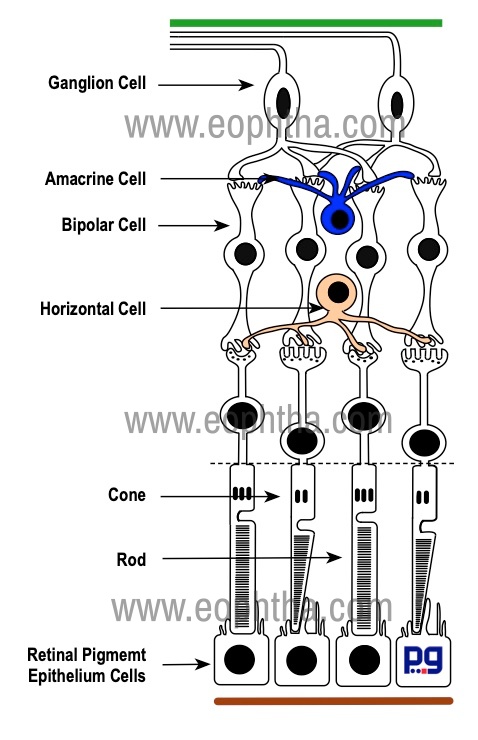
At optic disc only nerve fibers and astrocytes are present. Due to the absence of muller cells, this area is devoid of any internal limiting membrane. Anterior to the lamina cribrosa, the axons forming nerve fiber layer bundles are non-myelinated and become myelinated by oligodendrocytes once they cross the lamina cribrosa.
Near the ora serrata, the configuration of the retinal layer changes considerably. Here the inner and outer nuclear layers merge and there is a decrease in ganglion and nerve fiber layers, as well as photoreceptors (rods). The inner limiting membrane of the retina gradually coalesces with the vitreous base. The external limiting membrane continues into the ciliary body as a junctional zone between the pigmented and the nonpigmented epithelium. The rest of the residual sensory retina continues forward as the nonpigmented epithelium of the ciliary body. The RPE continues forward as the pigmented epithelium of the ciliary body.
|
Layers or Retina |
Common Characteristics |
Fovea/Macula |
|
RPE |
Retinal pigment epithelium cells; Single layer |
Present |
|
Layer of Photoreceptor |
Photoreceptors (Rods & Cones) |
Cones only |
|
Outer Nuclear |
Nuclei of Photoreceptors; 4 layers |
Present |
|
Outer Plexiform |
Receptor inner fibers, Bipolar and horizontal cell processes and synapses |
Present |
|
Inner Nuclear |
5-6 layers of cell nuclei |
Absent |
|
Inner Plexiform |
Present |
Absent |
|
Ganglion Cell Layer |
2-10 layers of nuclei |
In foveola, ganglion cell layer is absent; In fovea surrounding the foveola, it is 2-6 layers thick |
|
Nerve Fibre Layer |
Axons of ganglion cells |
Absent |
|
Inner Limiting Membrane |
Fusion of Muller cell and vitreous membranes |
Present, but very thin |
Blood Supply
Retina receives a dual circulation. Outer retinal layers derived blood supply from choroidal circulation whereas inner layers are dependent on post retinal blood vessels of the central retinal vessels. The retinal circulation exclusively supplies retina up to the inner nuclear layer. The outer plexiform layer is thought to receive its nutrients from both retinal and choroidal vessels.
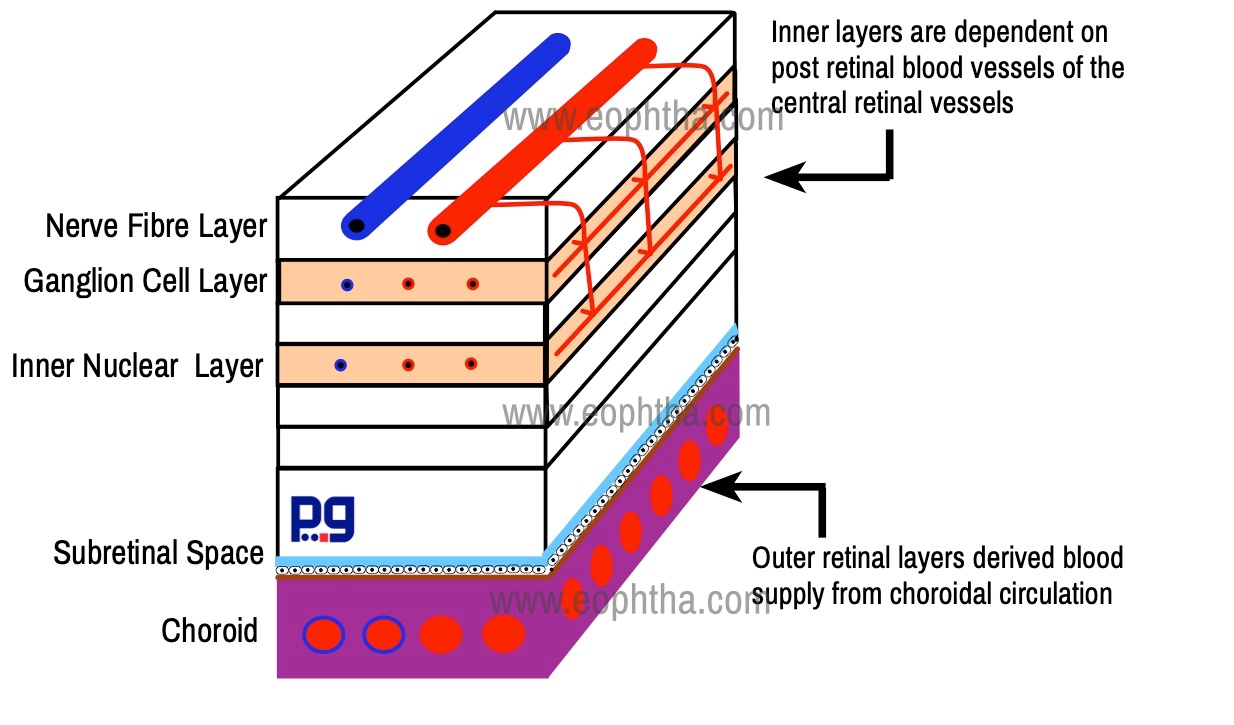
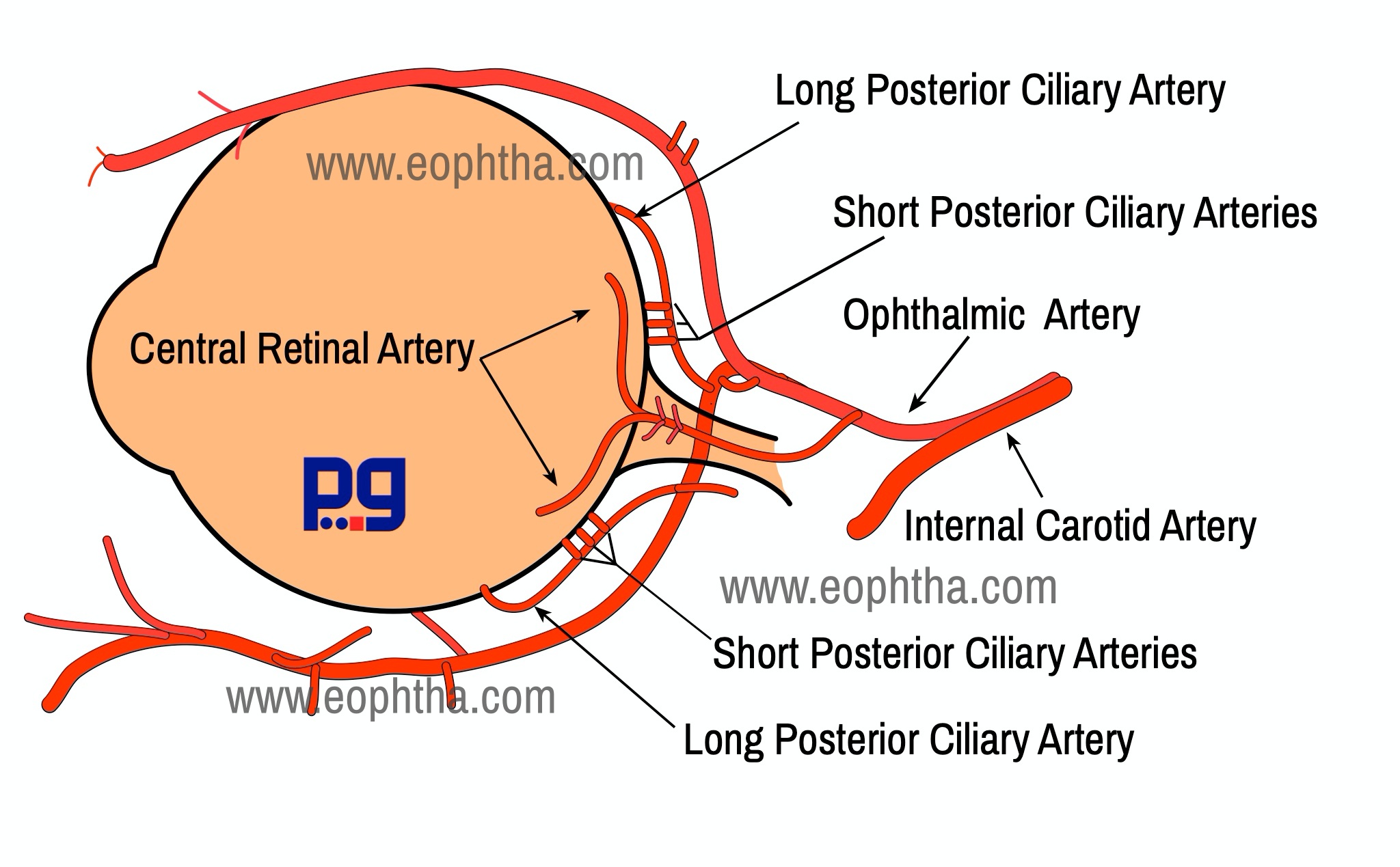
The central retinal artery, a branch of the ophthalmic artery, enters the retina through the optic disc. It divides into 4 main branches the upper and lower nasal and the upper and lower temporal. Retinal vessels are located in the nerve fiber layer just below the transparent internal limiting membrane. However, there are two types of capillary networks found in retina - superficial network is in the nerve fiber layer or ganglion cell layer and the deeper one is situated in the inner nuclear layer near the outer plexiform layer.
The central retinal artery inside the optic nerve has a well-developed tunica media which consists of 6 layer smooth muscle cells. While coming out from the optic nerve the muscular sheathing of the artery is replaced by an incomplete layer of pericytes. Pericytes are specialized mural cell with a contractile function that facilitates blood flow. Thus, histologically the central retinal vessel remains artery and vein only for a short distance found the optic disc, and turns into arterioles and venules. However, we use the term artery and vein both clinically and histologically.
Retinal vessels have no internal elastic lamina and are thus not affected in temporal arteritis
As the central nervous system, the retinal vessels are end vessels and normally do not anastomosis. However, anastomosis between the retinal vessels and the ciliary system of vessels occurs with the vessels which enter the optic nerve head from the circle of Zinn or Haler near the lamina cribrosa. This arterial circle is formed by an anastomosis between 2 or 4 or more short posterior ciliary arteries & lies in the sclera around the optic nerve.
Retinal vessels are the only vessels in the body that are not controlled by the autonomic nervous system. Autoregulation of retinal blood flow is mediated through smooth muscles of blood vessels and the number of vasoactive mediators secreted by the vascular endothelium. Increased oxygen tension causes vasoconstriction while increased intraocular pressure causes vasodilatation.
Also similar to the central nervous system vasculature The retinal blood vessels are lined by and endothelium with very tight cellular junctions. The pericytes and endothelial cells in retinal capillaries are present in a 1:1 ratio. The endothelial cells form a part of the blood-retinal barrier because they are joined by zonula occludens.
There are certain capillary-free zones in the retina. These are:
- Foveal Avascular Zone: Within the fovea is a region devoid of retinal blood vessels known as the foveal avascular zone
- An area of approximately 0.5 mm in diameter around all retinal vessels
- There is a capillary free zone in the retinal periphery situated approximately 1 mm near the ora serrata .
Retinal Vasculature: At a glance
Retinal Blood flow is 35-80μL/min.
Retina gets 4% of the total blood supply to the eye (choroid 85%)
Retinal arteries are narrower than veins
The Wall/lumen ratio of the artery is about 1:5 & in vein 1:10.
Retinal vessels are wider in adults than in children.
The arteriovenous ratio is 2:3.
Histology:
Artery = A thin tunica intima (an internal elastic lamina & a single layer of endothelial cells)* + tunica media (six layers of smooth muscles) + tunica adventitia
Vein = thinner than Arteries without Muscular layer
*it disappears in Lamina cribrosa & is absent in retinal arteries
The retina has the highest endothelial cell-to-pericyte ratio in the body, 1 : 1. 1
Reference:
- Sadda, S. R., Hinton, D. R., Schachat, A. P., Wilkinson, C. P., & Weidemann, P. (2018). Anatomy and Physiology. In Ryan's retina (pp. 1093-2595). New York, NY: Elsevier.
- Kolb H. Facts and Figures Concerning the Human Retina. In: Kolb H, Fernandez E, Nelson R, eds. Webvision: The Organization of the Retina and Visual System. University of Utah Health Sciences Center; 1995. Accessed August 19, 2020. http://www.ncbi.nlm.nih.gov/books/NBK11556/
- Masland RH. The Neuronal Organization of the Retina. Neuron. 2012;76(2):266-280. doi:10.1016/j.neuron.2012.10.002
- Foster, C. Stephen., Dimitri T. Azar, and Claes H. Dohlman. The Anatomy And Cell Biology Of The Human Cornea, Limbus, Conjunctiva, And Adnexa. In Smolin and Thoft's The Cornea: Scientific Foundations and Clinical Practice. Philadelphia: Wolters Kluwer, 2015.
- Remington, L. A., & Remington, L. A. (2012). Clinical anatomy and physiology of the visual system. Oxford: Elsevier Butterworth-Heinemann.
- Forrester, J. V. (2016). The eye: Basic sciences in practice. Edinburgh, London: Elsevier.
- Snell, R. S., & Lemp, M. A. (1998). Clinical anatomy of the eye. Oxford: Blackwell Science.